Del:



Veje af Pentose Faser og beslægtede sygdomme
den stien af pentoserne phosphat, også kendt som videresendelse monophosphat hexoser, er en grundlæggende metaboliske vej, hvis endelige produkt ribosen er nødvendig for syntesevejene af nukleotider og nukleinsyrer, såsom DNA, RNA, ATP, NADH, FAD og coenzym A.
Det producerer også NADPH (nikotinamid adenin dinucleotidphosphat), der anvendes i forskellige enzymatiske reaktioner. Denne rute er meget dynamisk og i stand til at tilpasse sine produkter afhængigt af de øjeblikkelige behov hos cellerne.
ATP (adenosintrifosfat) betragtes som "energivaluta" for cellen, fordi dens hydrolyse kan kobles til en bred vifte af biokemiske reaktioner.
Ligeledes bliver et andet NADPH kræves til reduktiv syntese af fedtsyrer, cholesterolsyntese, neurotransmittersyntese, fotosyntese og afgiftning reaktioner energi valuta, herunder.
Selv om NADPH og NADH er ens i struktur, kan de ikke anvendes udveksling i biokemiske reaktioner. NADPH deltager i brugen af fri energi ved oxidation af visse metabolitter til reduktiv biosyntese.
I modsætning hertil er NADH involveret i brugen af fri energi fra oxidation af metabolitter til syntetisering af ATP.
indeks
- 1 Historie og placering
- 2 funktioner
- 3 faser
- 3.1 Oxiderende fase
- 3.2 Ikke-oxidativ fase
- 4 relaterede sygdomme
- 5 referencer
Historie og beliggenhed
Indikationerne for eksistensen af denne rute begyndte i år 1930 takket være efterforskeren Otto Warburg, til hvem opdagelsen af NADP er tilskrevet ham+.
Visse observationer tillod opdagelsen af ruten, især fortsættelse af respiration i nærvær af glycolys inhibitorer, såsom fluoridionen.
Derefter beskriver forskerne Frank Dickens, Bernard Horecker, Fritz Lipmann og Efraim Racker i 1950 pentosephosphatvejen.
Vævene, der er involveret i syntesen af kolesterol og fedtsyrer, såsom brystkirtler, fedtvæv og nyrer, har høje koncentrationer af pentosephosphat enzymer.
Leveren er også et vigtigt væv til denne vej: Ca. 30% af oxidationen af glucose i dette væv sker takket være enzymerne af pentosephosphatvejen.
funktioner
Pentosephosphatvejen er ansvarlig for at opretholde carbonens homeostase i cellen. På samme måde syntetiserer vejen prækursorerne af nukleotider og molekyler involveret i syntese af aminosyrer (strukturelle blokke af peptider og proteiner).
Det er den vigtigste kilde til at reducere strømmen til enzymatiske reaktioner. Derudover giver den de nødvendige molekyler til anabolske reaktioner og til forsvarsprocesser mod oxidativ stress. Den sidste fase af sporet er kritisk i redox processer under stress situationer.
faser
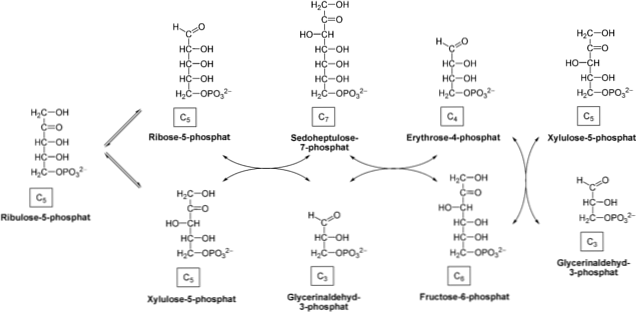
Pentosephosphatvejen består af to faser i cellecytosolen: oxidativ, NADPH genererer oxidationen af glucose-6-phosphat til ribose-5-phosphat; og en ikke-oxidativ, der involverer interomdannelse af sukkerarter tre, fire, fem, seks og syv carbonatomer.
Denne rute præsenterer delte reaktioner med Calvin-cyklen og med Entner-Doudoroff-banen, som er et alternativ til glycolyse.
Oxidativ fase
Oxidativ dehydrogenering fase begynder med et molekyle glucose-6-phosphat på carbon 1. Denne reaktion katalyseres af enzymet glucose-6-phosphatdehydrogenase, som har en høj specificitet for NADP+.
Produktet af denne reaktion er 6-phosphonoglucono-5-lacton. Derefter hydrolyseres dette produkt af enzymet lactonase for at give 6-phosphogluconat. Sidstnævnte forbindelse tages af enzymet 6-phosphogluconat-dehydrogenase og bliver ribulose-5-phosphat.
Fosfopentosa isomerase enzym katalyserer det sidste trin i den oxidative fase, der involverer syntesen af ribose-5-phosphat ved isomerisering af ribulose-5-phosphat.
Denne række reaktioner producerer to molekyler NADPH og et molekyle ribose 5-phosphat pr. Molekyle glucose 6-phosphat, der kommer ind i denne enzymatiske vej.
I nogle celler, NADPH krav er større end dem af ribose-5-phosphat. Derfor, transketolase og transaldolase enzymerne tage ribose-5-phosphat og konverterer glyceraldehyd 3-phosphat og fructose-6-phosphat, hvilket fører til ikke-oxidativ fase. Disse sidste to forbindelser kan komme ind i den glycolytiske vej.

Ikke-oxidativ fase
Fasen begynder med en epimeriseringsreaktion katalyseret af enzymet pentose-5-phosphat-epimerase. Ribulose-5-phosphat tages af dette enzym og omdannes til xylulose-5-phosphat.
Produktet optages af transketolase enzym handler sammen med coenzym thiaminpyrophosphat (TPP), der katalyserer passagen af xylulose-5-phosphat til ribose-5-phosphat. Ved overførsel af ketose til aldose produceres glyceraldehyd-3-phosphat og sedoheptulose-7-phosphat.
Derefter overfører transaldolase enzym C3 molekyle sedoheptulose-7-phosphat til glyceraldehyd-3-phosphat, som frembringer en fire-carbon sukker (erythrose-4-phosphat) og en seks-carbon (fructose-6 -phosphat). Disse produkter er i stand til at fodre glycolytic pathway.
Enzymet virker transcetosala igen at overføre en C2 xylulose 5-phosphat til erythrose-4-phosphat, hvilket fører til fructose-6-phosphat og resultere glyceraldehyd-3-phosphat. Som i det foregående trin kan disse produkter indgå i glycolyse.
Denne anden fase forbinder de veje, der genererer NADPH med dem, der er ansvarlige for syntetisering af ATP og NADH. Derudover kan produkterne fructose-6-phosphat og glyceraldehyd-3-phosphat indtaste gluconeogenese.
Relaterede sygdomme
Forskellige patologier er relateret til pentosephosphatets vej, mellem disse neuromuskulære sygdomme og forskellige typer kræft.
De fleste kliniske undersøgelser fokuserer på kvantificering af aktiviteten af glucose-6-phosphat dehydrogenase, fordi det er det primære enzym, der er ansvarlig for regulering af vejen.
I blodcellerne tilhørende personer, der er modtagelige for anæmi, har de en lav enzymatisk aktivitet af glucose-6-phosphat dehydrogenase. I modsætning hertil udviser cellelinjer relateret til carcinomer i strubehovedet høj enzymaktivitet.
NADPH involveret i produktionen af glutathion, en nøgle peptidmolekyle i beskyttelsen mod reaktive oxygenformer er involveret i oxidativt stress.
Cancere fører til aktiveringen af pentose og processer forbundet med metastase, angiogenese og reaktioner på behandlinger af kemoterapi og strålebehandling.
På den anden side udvikler kronisk granulomatøs sygdom, når der er mangel på produktion af NADPH.
referencer
- Berg, J.M., Tymoczko, J. L., Stryer, L (2002). Biokemi. WH Freeman
- Konagaya, M., Konagaya, Y., Horikawa, H., & Iida, M. (1990). Pentosephosphatvej i neuromuskulære sygdomme-evaluering af muskelglucose 6-phosphat dehydrogenase aktivitet og RNA indhold. Rinsho shinkeigak. Klinisk neurologi, 30(10), 1078-1083.
- Kowalik, M. A., Columbano, A., & Perra, A. (2017). Fremkaldende rolle af pentosephosphatvejen i hepatocellulær carcinom. Grænser i onkologi, 7, 87.
- Patra, K. C., & Hay, N. (2014). Pentosephosphatvejen og kræft. Udvikling i biokemiske videnskaber, 39(8), 347-354.
- Stincone, A., Prigione, A., Cramer, T., Wamelink, M., Campbell, K., Cheung, E., ... & Keller, M. A. (2015). Retur af metabolisme: biokemi og fysiologi af pentosephosphatvejen. Biologiske anmeldelser, 90(3), 927-963.
- Voet, D., & Voet, J. G. (2013). biokemi. Artmed Publisher.