Del:



Hvad er replikationsgaffelen?
den replikationsgaffel det er det punkt, hvor DNA replikation forekommer, det kaldes også et vækstpunkt. Det har en Y-form, og efterhånden som replikationen fortsætter, bliver hårnålen forskudt af DNA-molekylet.
DNA-replikation er den cellulære proces, der involverer duplikering af genetisk materiale i cellen. Strukturen af DNA er en dobbelt helix, og for at replikere dens indhold skal den åbnes. Hver af tråderne vil være en del af den nye DNA-kæde, da replikation er en semiconservativ proces.
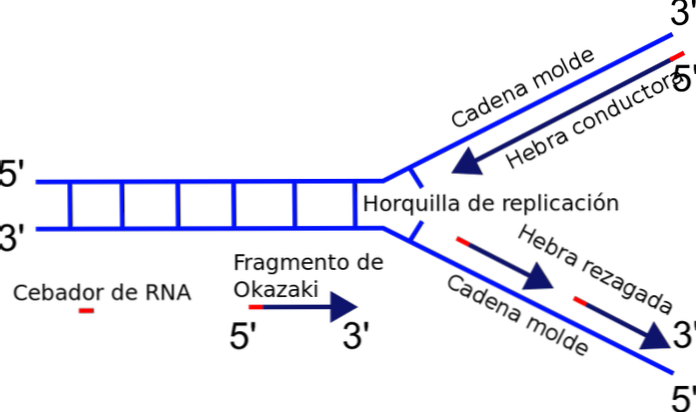
Replikationsgaffelen er dannet lige mellem krydset mellem de nyligt adskilte skabeloner eller skabelonkæder og det duplex-DNA, som endnu ikke er blevet dupliceret. Når man initierer DNA-replikation, kan en af strengene let duplikeres, mens den anden streng står over for et polaritetsproblem.
Det enzym, der er ansvarlig for polymerisering af kæden - DNA-polymerasen - syntetiserer kun DNA-strengen i 5'-3'-retningen. Således er en streng kontinuerlig, og den anden lider en diskontinuerlig replikation, der frembringer fragmenter af Okazaki.
indeks
- 1 Replikation af DNA og replikationsgaffel
- 1.1 Envejs- og tovejsreplikation
- 1.2 Enzymer involveret
- 1.3 Start af replikation og gaffeldannelse
- 1.4 Forlængelse og bevægelse af gaflen
- 1.5 Afslutning
- 2 DNA-replikation er semikonservativ
- 3 Polaritetsproblemet
- 3.1 Hvordan polymerasen virker?
- 3.2 Produktion af Okazaki-fragmenter
- 4 referencer
Replikation af DNA og replikationsgaffel
DNA er molekylet, der indeholder de nødvendige genetiske oplysninger af alle levende organismer - med undtagelse af nogle vira.
Denne enorme polymer sammensat af fire forskellige nukleotider (A, T, G og C) ligger i kernen af eukaryoter i hver af de celler, der udgør vævene af disse væsener (undtagen i modne røde blodlegemer af pattedyr, der mangler af kernen).
Hver gang en celle opdeles, skal DNA'et replikeres for at stamme fra en dattercelle med genetisk materiale.
Envejs- og tovejsreplikation
Replication kan være ensrettet eller tovejs, afhængigt af dannelsen af replikationsgaffeln ved oprindelsesstedet.
Logisk er der i tilfælde af replikation i én retning kun dannet en gaffel, mens tovejsreformation i tovejsreplikation dannes.
Enzymer involveret
Til denne proces er der brug for et komplekst enzymatisk maskineri, der fungerer hurtigt, og som kan replikere DNA'et på en præcis måde. De vigtigste enzymer er DNA-polymerase, DNA-primase, DNA-helicase, DNA-ligase og topoisomerase.
Start af replikation og dannelse af gaffelen
DNA-replikation starter ikke på et vilkårligt sted i molekylet. Der er specifikke regioner i DNA'et, der markerer begyndelsen af replikationen.
I de fleste bakterier har bakteriekromosomet et enkelt AT-rigt udgangspunkt. Denne sammensætning er logisk, da det letter åbningen af regionen (AT-parene er forbundet med to brintbroer, mens GC-parret med tre).
Når DNA'et begynder at åbne, dannes en Y-formet struktur: replikationsgaflen.
Forlængelse og bevægelse af gaflen
DNA-polymerase kan ikke starte syntesen af datterkæder fra bunden. Du har brug for et molekyle, der har en 3'-ende, så polymerasen har, hvor man begynder at polymerisere.
Denne gratis 3'-ende tilbydes af et lille molekyle nukleotider kaldet primer eller primer. Den første virker som en slags krog for polymerasen.
Med replikationsforløbet har replikationsgaflen evnen til at bevæge sig langs DNA'et. Pladsen af replikationsgaflen forlader to enkeltbånds-DNA-molekyler, der dirigerer dannelsen af dobbeltbånds datter molekyler.
Gaffelen kan forøges takket være virkningen af helikaseenzymerne, der afvikler DNA-molekylet. Dette enzym bryder hydrogenbindingerne mellem basisparene og tillader forskydningen af gaflen.
færdiggørelse
Replikationen afsluttes, når de to gafler er ved 180 ° C fra oprindelsen.
I dette tilfælde taler vi om, hvordan replikationsprocessen i bakterierne flyder, og det er nødvendigt at fremhæve hele torsionsprocessen i det cirkulære molekyle, der involverer replikation. Topoisomeraser spiller en vigtig rolle i afviklingen af molekylet.
DNA-replikation er semikonservativ
Har du nogensinde spekuleret på, hvordan replikation forekommer i DNA? Det vil sige, at der skal opstå en anden dobbelt helix fra den dobbelte helix, men hvordan sker det? I flere år var dette et åbent spørgsmål blandt biologer. Der kunne være flere permutationer: to gamle tråde sammen og to nye sammen, eller en ny tråd og en gammel til at danne dobbelt spiral.
I 1957 blev dette spørgsmål besvaret af forskere Matthew Meselson og Franklin Stahl. Replikationsmodellen foreslået af forfatterne var den semikonservative.
Meselson og Stahl udtalte, at resultatet af replikation er to dobbeltstrengede DNA-molekyler. Hvert af de resulterende molekyler er sammensat af en gammel streng (fra moderen eller det indledende molekyle) og en nyligt syntetiseret ny streng..
Polaritetsproblemet
Hvordan polymerase virker?
DNA-helixen er dannet af to kæder, der løber på en antiparallel måde: den ene går i 5'-3'-retningen og en anden 3'-5 '.
Det vigtigste enzym i replikationsprocessen er DNA-polymerase, som er ansvarlig for at katalysere bindingen af de nye nucleotider, som vil blive tilsat til kæden. DNA-polymerasen kan kun forlænge kæden i 5'-3'-retningen. Denne kendsgerning forhindrer samtidig duplikering af kæderne i replikationsgaflen.
Hvorfor? Tilsætningen af nukleotider forekommer ved den frie ende 3 'hvor der findes en hydroxylgruppe (-OH). Således kan kun en af kæderne let forstærkes ved den terminale tilsætning af nukleotidet til 3'-enden. Dette kaldes ledende eller kontinuerlig streng.
Fremstilling af Okazaki-fragmenter
Den anden streng kan ikke forlenge, fordi den frie ende er 5 'og ikke 3', og ingen polymerase katalyserer tilsætningen af nukleotider til 5'-enden. Problemet løses ved syntese af flere korte fragmenter (130 til 200 nukleotider), hver i den normale replikationsretning fra 5 'til 3'.
Denne diskontinuerlige syntese af fragmenter slutter med foreningen af hver af delene, en reaktion katalyseret af DNA-ligase. Til ære for opdageren af denne mekanisme, Reiji Okazaki, kaldes de små segmenter, der er syntetiseret, fragmenter af Okazaki.
referencer
- Alberts, B., Bray, D., Hopkin, K., Johnson, A. D., Lewis, J., Raff, M., ... & Walter, P. (2015). Væsentlig cellebiologi. Garland Science.
- Cann, I. K., & Ishino, Y. (1999). Archaeal DNA replikation: identificere stykkerne for at løse et puslespil. Genetik, 152(4), 1249-67.
- Cooper, G. M., & Hausman, R. E. (2004). Cellen: Molekylær tilgang. Medicinska naklada.
- Garcia-Diaz, M. & Bebenek, K. (2007). Multiple funktioner af DNA-polymeraser. Kritiske anmeldelser i plantevidenskab, 26(2), 105-122.
- Lewin, B. (2008). gener IX. Mc Graw-Hill Interamericana.
- Shcherbakova, P.V., Bebenek, K., & Kunkel, T. A. (2003). Funktioner af eukaryotiske DNA-polymeraser. Videnskabens SAGE KE, 2003(8), 3.
- Steitz, T. A. (1999). DNA-polymeraser: strukturel mangfoldighed og fælles mekanismer. Journal of Biological Chemistry, 274(25), 17395-17398.
- Watson, J. D. (2006). Molekylærbiologi af genet. Ed. Panamericana Medical.
- Wu, S., Beard, W. A., Pedersen, L.G., og Wilson, S.H. (2013). Strukturel sammenligning af DNA-polymerasarkitektur antyder en nukleotidgateway til det polymeraseaktive sted. Kemiske anmeldelser, 114(5), 2759-74.