Del:



Tubulina Alfa og Beta, Funktioner
den tubulin er et globulært dimerisk protein dannet af to polypeptider: tubulin alfa og beta. De er organiseret i form af et rør, der giver anledning til mikrotubuli, som sammen med actinmikrofilamenterne og de mellemliggende filamenter udgør cytoskelet.
Mikrotubuli findes i forskellige essentielle biologiske strukturer, såsom sædets flagellum, forlængelsen af de cilierede organismer, luftarterne i luftrøret og æggelederne, blandt andre..
Desuden er de strukturer, der danner tubulina-funktionen som transportruter, analoge til sporene af et spor af materialer og organeller inde i cellen. Fordelingen af stoffer og strukturer er mulig takket være motoriske proteiner forbundet med mikrotubuli kaldet kinesin og dynein.
indeks
- 1 Generelle egenskaber
- 2 tubulin alfa og beta
- 3 funktioner
- 3.1 Cytoskelet
- 3.2 Mitose
- 3.3 Centrosome
- 4 Evolutionært perspektiv
- 5 referencer
Generelle egenskaber
Tubulin-underenhederne er heterodimerer på 55.000 dalton og er byggestenene af mikrotubuli. Tubulin findes i alle eukaryote organismer og har været stærkt bevaret i løbet af evolutionen.
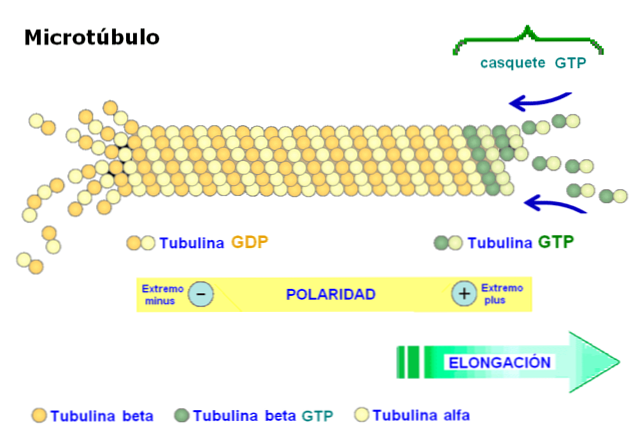
Dimeren er sammensat af to polypeptider kaldet tubulin alfa og beta. Disse polymeriseres til dannelse af mikrotubuli, som består af tretten protofilamenter arrangeret parallelt i form af et hul rør.
Et af de mest relevante egenskaber ved mikrotubuli er strukturens polaritet. Med andre ord er mikrotubulets to ender ikke ens: den ene ende hedder den hurtigt voksende ende eller "mere", og den anden ende er langsomt voksende eller "mindre"..
Polaritet er vigtig, da det bestemmer bevægelsesretningen langs mikrotubuli. Tubulin-dimeren er i stand til at polymerisere og depolarisere i hurtige samlingscykler. Dette fænomen forekommer også i actinfilamenter.
Der er en tredje type underenhed: det er gamma tubulin. Dette er ikke en del af mikrotubuli og er placeret i centrosomer; Det deltager dog i kernen og dannelsen af mikrotubuli.
Tubulin alfa og beta
Alfa- og beta-underenhederne er stærkt forbundet med dannelse af en kompleks heterodimer. Faktisk er interaktionen af komplekset så intens, at det ikke adskiller under normale forhold.
Disse proteiner er dannet af 550 aminosyrer, mest syrer. Selv om alfa- og beta-tubulinerne er ret ens, kodes de af forskellige gener.
I tubulina alfa kan findes aminosyrerester med en acetylgruppe, der giver forskellige egenskaber i celle flagella.
Hver subunit af tubulin er forbundet med to molekyler i alfa-tubulin GTP binder irreversibelt og hydrolyse af forbindelsen ikke forekommer, mens den anden bindingssted på beta tubulin reversibelt binder GTP og hydrolyserer.
GTP hydrolyse resulterer i et fænomen kaldet "dynamisk ustabilitet", hvor mikrotubuli undergår cykler af stigning og fald, afhængigt af hastigheden for tilsætning af tubulin og hastigheden for hydrolyse af GTP.
Dette fænomen betyder en høj omsætningshastighed for mikrotubuli, hvor halveringstiden for strukturen er kun få minutter.
funktioner
cytoskelet
Alfa- og beta-underenhederne af tubulin polymeriserer for at give anledning til mikrotubuli, som er en del af cytoskelettet.
Ud over mikrotubuli består cytoskelettet af to yderligere strukturelle elementer: actinmikrofilamenterne på ca. 7 nm og de mellemliggende filamenter med en diameter på 10 til 15 nm.
Cytoskelettet er rammen af cellen, giver den støtte og opretholder den cellulære form. Imidlertid er membranen og de subcellulære rum ikke statiske og er i konstante bevægelser for at kunne udføre fænomenerne endocytose, fagocytose og sekretion af materialer.
Ciroskelets struktur gør det muligt for cellen at rumme sig selv for at opfylde alle de nævnte funktioner.
Det er det ideelle medium til cellulære organeller, plasmamembranen og andre cellulære komponenter til at udføre deres sædvanlige funktioner, ud over at deltage i celledeling.
De bidrager også i fænomener af cellulære bevægelser som amoebas bevægelse og i specialiserede strukturer til forskydning som cilia og flagella. Endelig er det ansvarligt for bevægelsen af musklerne.
mitose
Takket være den dynamiske ustabilitet kan mikrotubuli omorganiseres fuldstændigt under celledeling. Mikrotubuleringsarrangementet under grænsefladen er i stand til at adskille og tubulinunderenhederne er frie.
Tubulin kan samles igen og stamme fra den mitotiske spindel, som deltager i adskillelsen af kromosomerne.
Der er visse lægemidler, såsom colchicin, taxol og vinblastin, der afbryder cellefordelingsprocesser. Virker direkte på tubulinmolekyler, der påvirker montering og dissociation fænomenet af mikrotubuli.
centrosom
I dyreceller, mikrotubuli forlænge centrosomet, en nær kernestruktur dannet af et par af centrioler (hver orienteret vinkelret) og omgivet af et amorft stof, kaldet pericentriolar matrix.
Centriolerne er cylindriske kroppe dannet af ni tripletter af mikrotubuli, i en organisation, der ligner cellulære cilia og flagella.
I processen med celledeling strækker mikrotubuli sig fra centrosomerne, der danner den mitotiske spindel, der er ansvarlig for den korrekte fordeling af kromosomerne til de nye datterceller..
Det lader til, at centrioler ikke er væsentlige for mikrotubulusaggregering i celler, fordi ikke præsentere i planteceller eller i visse eukaryote celler, såsom æg af visse gnavere.
I den pericentriolære matrix forekommer initiering til samling af mikrotubuli, hvor kernedannelse sker ved hjælp af gamma tubulin.
Evolutionært perspektiv
De tre typer af tubulin (alpha, beta og gamma) er kodet af forskellige gener og er homologe med et gen, der findes i prokaryoter, der koder for et protein på 40.000 dalton, kaldet FtsZ. Det bakterielle protein svarer funktionelt og strukturelt til tubulin.
Det er sandsynligt, at proteinet havde en forfædrefunktion i bakterierne og blev modificeret under de evolutionære processer og afsluttede i et protein med de funktioner, som det spiller i eukaryoter..
referencer
- Cardinali, D. P. (2007). Anvendt neurovidenskab: dens fundament. Ed. Panamericana Medical.
- Cooper, G.M. (2000). Cellen: En molekylær tilgang. 2. udgave. Sunderland (MA): Sinauer Associates.
- Curtis, H., & Schnek, A. (2006). Invitation til biologi. Ed. Panamericana Medical.
- Frixione, E., & Meza, I. (2017). Levende maskiner: Hvordan cellerne bevæger sig?. Økonomisk Kulturfond.
- Lodish H, Berk A, Zipursky SL, et al. (2000). Molecular Cell Biology. 4. udgave. New York: W. H. Freeman.