Del:



Unionsceller og deres egenskaber
den celleforbindelser de er de kontaktbroer der eksisterer mellem de cytoplasmatiske membraner mellem tilstødende celler eller mellem en celle og matrixen. Leddene afhænger af den undersøgte type væv, der fremhæver forbindelserne mellem epitel-, muskel- og nerveceller.
I cellerne er der molekyler relateret til adhæsionen mellem dem. Der er dog behov for yderligere elementer, der øger stabiliteten af bindingen af væv. Dette opnås med cellulære kryds.
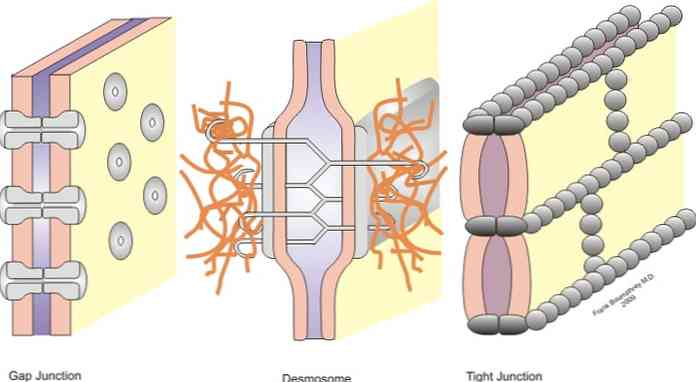
Leddene er klassificeret i symmetriske led (smalle led, desmosomer i bæltet og slidset led) og asymmetriske led (hemidesmosomer).
Smalle krydsninger, bælte desmosomer, punkt desmosomer og hæmomosmer er led, der tillader forankring; mens krydsningerne i spalten opfører sig som broer af forening mellem de nærliggende celler, hvilket tillader udveksling af opløste stoffer mellem cytoplasmaerne.
Bevægelsen af opløste stoffer, vand og ioner forekommer gennem og mellem individuelle cellulære komponenter. Således er der den transcellulære vej, der styres af en række ledninger og transportører. I modsætning til den paracellulære vej, som reguleres af kontakter mellem celler - det vil sige celleforbindelser.
I planter finder vi cellulære krydsninger, der ligner slidsforbindelser, kaldet plasmodesmermer. Selvom de er forskellige i deres struktur, er funktionen den samme.
Af medicinsk synspunkt resulterer visse mangler i celleforbindelser i erhvervede eller arvelige sygdomme forårsaget af skade på epitelbarrieren.
indeks
- 1 kendetegn
- 2 typer
- 2.1 - Smarte forbindelser
- 2.2 - Spalte eller mellemrumskryds
- 2.3 - Forankring eller vedhængende forbindelser
- 2.4 -Hemidesmosomer
- 3 Cellulære krydsninger i planter
- 4 Medicinsk perspektiv
- 5 referencer
funktioner
Levende organismer består af diskrete og varierede strukturer kaldet celler. Disse er afgrænset af en plasmamembran, som holder dem adskilt fra det ekstracellulære miljø.
Men selv om de er bestanddelene af levende væsener, ligner de ikke mursten, da de ikke er isoleret fra hinanden.
Cellerne er elementer, der kommunikeres med hinanden og med det ekstracellulære miljø. Derfor skal der være en måde for cellerne at danne væv og kommunikere, mens membranen forbliver intakt.
Dette problem er løst takket være forekomsten af celleforbindelser, der findes i epithelia. Disse krydsninger dannes mellem to tilstødende celler klassificeres efter funktionen af hver i symmetriske og asymmetriske ledd.
Asymmetriske forbindelser tilhører hemidesmosomer, og ved symmetriske forbindelser tætte forbindelser, et desmosom på et bælte, desmosomer og slidsforbindelser. Næste vil vi detaljeret beskrive hver af fagforeningerne.
typen
-Smalle kryds

Smalle krydsninger, der også er kendt i litteraturen som okklusive kryds, er sektorer i cellemembraner i naboceller, der er tæt forbundet - som navnet "smalt kryds" angiver.
Under gennemsnitlige betingelser adskilles cellerne med en afstand på 10 til 20 nm. I tilfælde af smalle krydsninger reduceres denne afstand imidlertid betydeligt, og begge cellers membraner berøres eller endog fusioneres.
Et typisk snævert kryds er placeret mellem sidevæggene i nabostationerne i en minimal afstand fra deres apikale overflader.
I epithelvævet etablerer alle celler en fagforening af denne type for at forblive forenet. I denne interaktion er cellerne placeret som et mønster der minder om en ring. Disse fagforeninger dækker hele omkredsen.
Proteiner involveret i tætte krydsninger
Ocludina og Claudina
Områderne med tæt kontakt omgiver hele overfladen af cellen. Disse regioner danner anastomoserende kontaktstrimler af transmembranproteinerne kendt som occludin og claudin. Udtrykket anastomose henviser til foreningen af visse anatomiske elementer.
Disse to proteiner tilhører gruppen af tetraespaniner. De er karakteriseret ved at have fire transmembrane domæner, to eksterne sløjfer og to relativt korte cytoplasmatiske haler.
Det er blevet observeret, at occludin interagerer med fire andre proteinmolekyler, kaldet occludin fra zonulaen og forkortet som ZO. Denne sidste gruppe omfatter proteiner ZO 1, ZO 2, ZO 3 og afuna.
Claudin er derimod en familie på 16 proteiner, der udgør en række lineære fibriller i de smalle kryds, hvilket gør det muligt for denne union at optræde som en "barriere" i den paracellulære vej..
Nectinas og JAM
Nektinerne og adhesionsmolekylerne i fagforeningerne (forkortet af dens akronym i engelsk JAM) fremgår også af de smalle kryds. Disse to molekyler findes som homodimerer i det intracellulære rum.
Nektinerne er forbundet med actinfilamenterne ved hjælp af afadinproteinet. Sidstnævnte ser ud til at være afgørende, da de i slettelsen af genet kodende for afadin hos gnavere fører til embryoens død.
Funktioner af smalle kryds
Disse typer af celle-til-celle krydsninger udfører to væsentlige funktioner. Den første er at bestemme polariteten af cellerne i epitelet, at adskille det apikale domæne fra basolateret og forhindre en unødig diffusion af lipider, proteiner og andre biomolekyler..
Som vi nævnte i definitionen, grupperes cellerne i epitelet i en ring. Denne struktur adskiller den apikale overflade af cellen fra de laterale og basale, som fastlægger differentieringen mellem domænerne.
Denne adskillelse betragtes som en af de vigtigste begreber i undersøgelsen af epithelys fysiologi.
For det andet forhindrer de stramme krydsninger fri passage af stoffer gennem laget af epithelceller, som omdannes til en barriere mod den paracellulære vej..
-Spalte eller mellemrumskryds

Slids- eller mellemkrydsninger findes i områder uden begrænsende cytoplasmisk membran mellem naboceller. I et splittet knudepunkt er cytoplasmaerne i cellerne forbundet, og der skabes en fysisk forbindelse, hvor små molekylers passage kan forekomme.
Denne klasse af led er fundet i stort set alle epithelia og i andre typer væv, hvor de tjener ganske varierede formål..
For eksempel kan slidsforbindelserne i flere væv åbne eller lukke som reaktion på ekstracellulære signaler, som det er tilfældet med neurotransmitterdopaminen. Tilstedeværelsen af dette molekyle reducerer kommunikationen mellem en klasse neuroner i nethinden som reaktion på den øgede intensitet af lys.
Proteiner involveret i spaltning leddene
Spalteforbindelser dannes af proteiner kaldet connexiner. Således opnås en "konexon" ved foreningen af seks connexin-monomer. Denne struktur er en hul cylinder, der passerer gennem den cytoplasmatiske membran.
Forbindelserne er arrangeret således, at der dannes en kanal mellem cytoplasmaerne hos de tilstødende celler. Derudover har connexons tendens til at aggregere og danne en slags plader.
Funktioner af slidsamlinger
Takket være dannelsen af disse bindinger kan bevægelsen af visse molekyler mellem de tilstødende celler forekomme. Størrelsen af molekylet, der skal transporteres, er afgørende, den optimale diameter er 1,2, såsom calciumioner og cyclisk adenosinmonophosphat.
Specifikt er det de uorganiske ioner og de vandopløselige molekyler, som kan overføres fra en cellulær cytoplasma til den tilstødende cytoplasma.
Calciumkoncentrationer spiller en afgørende rolle i denne kanal. Når calciumkoncentrationen stiger, har de aksiale kanaler tendens til at lukke.
På denne måde deltager slidsledene aktivt i processen med elektrisk og kemisk kobling mellem celler, som forekommer i hjertemuskelcellerne, som er ansvarlige for transmission af elektriske impulser.
-Forankring eller vedhængende forbindelser

Under de smalle led, finder vi forankringsledene. Generelt er disse placeret i nærheden af epitelens apikale overflade. I denne gruppe kan vi skelne mellem tre hovedgrupper, zonula adherens eller desmosoma i bæltet, macula adherens eller desmosome punkt og desmosome.
I denne type kryds er adskilte cellemembraner, der er forbundet med zonules og adherens macules, adskilt af en cellulær afstand, der er relativt bred - hvis vi sammenligner dem med det mindste rum, der eksisterer i tilfælde af smalle kryds..
Det intercellulære rum er optaget af proteiner, der tilhører familien af cadheriner, desmogleiner og desmocoliner forbundet med cytoplasmatiske plader, der har andre proteiner kaldet desmoplaquina, placoglobina og placofilina.
Klassificering af forankringsled
Zonula adherens
Som i tilfælde af smalle led, observerer vi også arrangementets mønster i form af en ring eller et bælte. Zonula-adherens er forbundet med actinmikrofilm gennem interaktion mellem to proteiner: cadheriner og cateniner.
Macula adherens
I nogle tilfælde er denne struktur simpelthen kendt som desmosom, det er et punkteret kryds, der er forbundet med mellemliggende filamenter dannet af keratin. I denne sammenhæng kaldes de nævnte keratinstrukturer "tonofilimanetos". Filamenterne strækker sig fra et punkt til et andet i epithelcellerne.
Point desmosomas
Disse giver styrke og stivhed til epithelcellerne. Således antages det, at deres primære funktion er relateret til styrkelse og stabilisering af tilstødende celler.
Desmosomer kan sammenlignes med en slags nitter eller svejsninger, da de ligner separate små pletter og ikke kontinuerlige bånd.
Vi finder denne type led i de interkalierede diske, som forbinder kardiocytterne i hjertemusklen og i meningerne, der dækker den ydre overflade af hjernen og rygmarven..
-hemidesmosome

Hemidesmosomer falder ind i kategorien af asymmetriske kryds. Disse strukturer har funktionen at forankre epitelcelleens basale domæne med den underliggende basale lamina.
Betegnelsen hemidesmosom anvendes fordi denne struktur virker, bogstaveligt talt "medium" desmosoma. Men ud fra deres biokemiske sammensætning er begge fagforeninger helt forskellige.
Det er vigtigt at præcisere, at desmosomerne er ansvarlige for at adhærere en nabocelle med en anden, mens funktionen af hemidesmosomet er at slutte sig til cellen med den basale lamina.
Modsætning adherens macula eller desmosom, hemidesmosomer har en anden struktur, der består af: et cytoplasmatisk ark forbundet med intermediære filamenter og plade ydre membraner, som er ansvarlig for fastgørelse af hemidesmosome med den basale lamina, gennem en ankerfilament.
En af funktionerne i hæmosmosomer er at øge den generelle stabilitet af epitelvæv, takket være tilstedeværelsen af intermediære filamenter i cytoskelettet, der er fastgjort til bestanddelene i basal laminatet.
Cellforbindelser i planter
Planteriget mangler de fleste af de ovenfor nævnte celleforbindelser med undtagelse af en funktionel modstykke, der ligner slidsforbindelser.
I planter er cytoplasmaerne hos tilstødende celler forbundet med veje eller kanaler kaldet plasmododer.
Denne struktur skaber et kontinuum fra en plantecelle til den næste. Skønt det adskiller sig strukturelt fra spaltekrydsene, har de meget lignende roller, der tillader passage af små ioner og molekyler.
Medicinsk perspektiv
Fra medicinens synspunkt er celleforeninger et relevant spørgsmål. Det har vist sig, at mutationer i generne, der koder for de proteiner, der er involveret i krydsene, oversættes til kliniske patologier.
For eksempel, hvis der er en vis mutation i genet, der koder for en bestemt type claudin (et af de proteiner, der formidler interaktionen i smalle type kryds), forårsager det en sjælden sygdom hos mennesker..
Dette er syndromet af tab af renal magnesium, og symptomerne omfatter en lav mængde magnesium og anfald.
Derudover er det blevet konstateret, at en mutation i genet, som koder for protein nektin 1, er ansvarlig for spaltpalatsyndrom eller spaltelæbe. Denne tilstand betragtes som en af de mest almindelige misdannelser hos nyfødte.
Mutationer i nektin 1-genet har også været forbundet med en anden tilstand, der hedder ektodermal dysplasi, der påvirker menneskers hud, hår, negle og tænder.
Foliar pemphigus er en bullous patologi af huden bestemt af autoantistoffer mod desmoglein 1, et nøgleelement der er ansvarlig for at opretholde sammenhængen i epidermis.
referencer
- Alberts, B., Bray, D., Hopkin, K., Johnson, A. D., Lewis, J., Raff, M., ... & Walter, P. (2015). Væsentlig cellebiologi. Garland Science.
- Cooper, G. M., & Hausman, R. E. (2000). Cellen: Molekylær tilgang. Sinauer Associates.
- Curtis, H., & Barnes, N. S. (1994). Invitation til biologi. Macmillan.
- Hill, R. W., Wyse, G. A., Anderson, M. & Anderson, M. (2004). Dyrefysiologi. Sinauer Associates.
- Karp, G. (2009). Celle- og molekylærbiologi: begreber og eksperimenter. John Wiley & Sons.
- Kierszenbaum, A., & Tres, L. (2016). Histologi og cellebiologi: en introduktion til patologi. Elsevier Brasilien.
- Lodish, H., Berk, A., Darnell, J. E., Kaiser, C. A., Krieger, M., Scott, M.P., ... & Matsudaira, P. (2008). Molekylcellebiologi. Macmillan.
- Voet, D., & Voet, J. G. (2006). biokemi. Ed. Panamericana Medical.